Содержание
Регулировка створок — СТВОРКА Окна | Балконы
Регулировка створок ПВХ окон
Регулировка пластиковых окон — это одно из многих достоинств относительно старых деревянных окон. В процессе эксплуатации окна работа створок может быть нарушена. Это может быть вызвано разными причинами, о которых мы Вам тоже расскажем. Главное не паниковать! Регулировка окон может быть произведена своими руками или при помощи специалиста компании в рамках гарантийного обслуживания. Современная оконная фурнитура имеет несколько регулировочных винтов, которые позволяют привести створку в нормальное рабочее состояние.
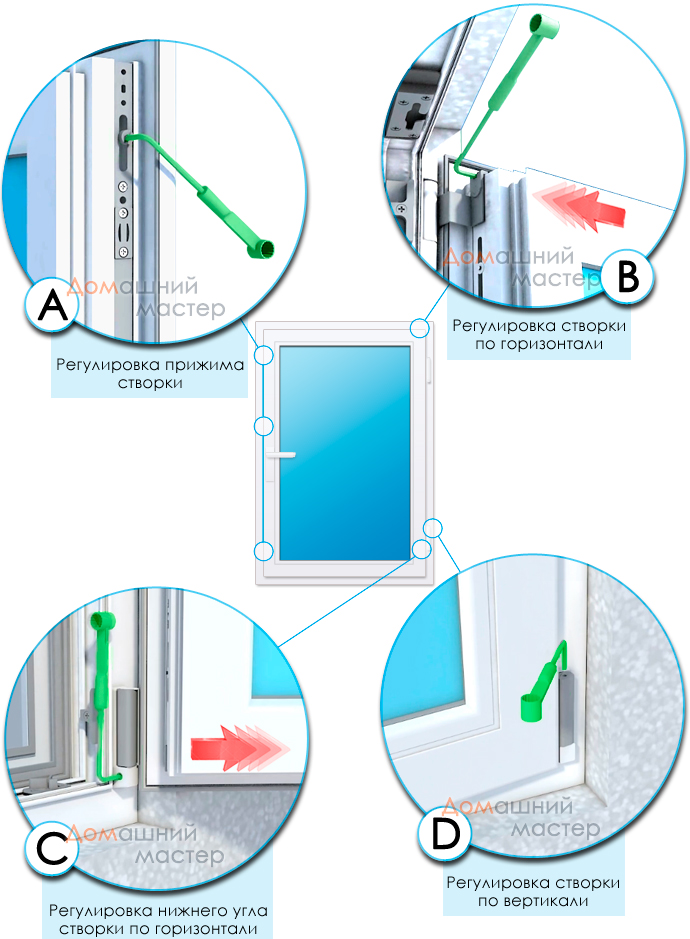
Винт № 1. Находится в нижней петле створки.
Этот винт позволяет производить регулировку створки по высоте, т.е. поднимать ее и опускать. Чтобы добраться к нему, необходимо снять декоративную накладку петли. Вращением винта по часовой стрелке или против часовой стрелки поднимаем или опускаем створку. В зависимости от производителя фурнитуры винты могут иметь различные формы под разный инструмент. Чаще всего регулировка производится шестигранником №4. Встречаются винты под шестигранники меньших размеров, а также винты «под звездочку» (как у фурнитуры G-U). Очень редко можно встретить совсем уж экзотические решения в виде фигурных гаек (фурнитура «AUBI» старого образца или современная Siegenia-Aubi Titan AF).
Чаще всего регулировка производится шестигранником №4. Встречаются винты под шестигранники меньших размеров, а также винты «под звездочку» (как у фурнитуры G-U). Очень редко можно встретить совсем уж экзотические решения в виде фигурных гаек (фурнитура «AUBI» старого образца или современная Siegenia-Aubi Titan AF).
Для створок с правым открыванием: вращение по часовой стрелке поднимает створку, против часовой стрелки — опускает. Для створок с левым открыванием соответственно наоборот: вращение по часовой стрелке опускает створку, против часовой стрелки — поднимает.
Винт № 2. Находится на опоре нижней петли.
Этот винт позволяет производить регулировку створки по ширине, т.е. перемещать ее влево-вправо. Чтобы добраться к нему, необходимо снять декоративную накладку петли. Впрочем, большинство декоративных накладок имеют специальное окошко для доступа к этому винту. Вращением винта по часовой стрелке или против часовой стрелки перемещаем створку влево или вправо. В зависимости от производителя фурнитуры эти винты также могут иметь различные формы под разный инструмент. Чаще всего регулировка производится шестигранником №4. Встречаются винты под шестигранники меньших размеров, а также винты «под звездочку».
В зависимости от производителя фурнитуры эти винты также могут иметь различные формы под разный инструмент. Чаще всего регулировка производится шестигранником №4. Встречаются винты под шестигранники меньших размеров, а также винты «под звездочку».
ВНИМАНИЕ! Направление вращения винта определяется в зависимости от того, с какой стороны окошка петли помещается регулировочный ключ. Обычно проще определить направление вращения экспериментальным путем)).
Винт № 3. Находится на верхней петле
Этот винт также, как и винт № 2, позволяет производить регулировку створки по ширине, перемещать ее влево-вправо, но используется гораздо чаще. Это обусловлено тем, что вращением этого винта можно приподнимать или опускать угол створки с противоположной от петель стороны. Актуально при провисании створок.
Для створок с правым открыванием: вращение по часовой стрелке поднимает угол створки, против часовой стрелки — опускает. Для створок с левым открыванием соответственно наоборот: вращение по часовой стрелке опускает угол створки, против часовой стрелки — поднимает.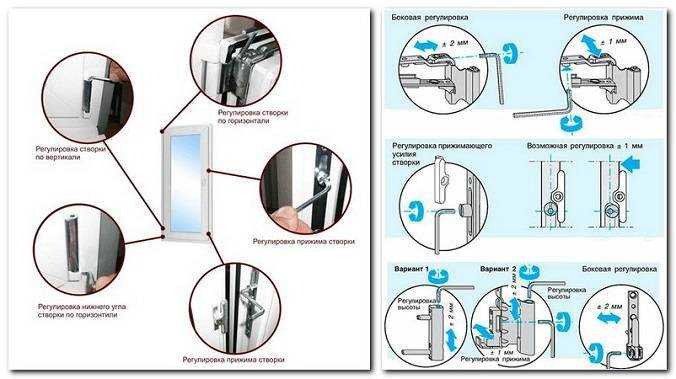
Регулировка прижима створки
Это основные регулировочные винты, которые позволяют изменять положение створки относительно рамы окна. Некоторые нижние петли створок также имеют дополнительные регулировочные винты, позволяющие менять степень прижима в районе нижней петли. Также имеются регулировочные винты в цапфах приводов и угловых передач. Они служат для изменения степени прижима створки к раме. Это происходит благодаря тому, что цапфа имеет форму эксцентрика, и, изменяя ее положение относительно своей оси, изменяется давление цапфы на запорную пластину, а следовательно и прижим. Цапфа имеет отметку (риску), по которой можно определить, в каком именно режиме находится ее работа. Если риска смотрит вдоль фурнитурного паза — это среднее положение, установлено по умолчанию на новой фурнитуре. Если риска смотрит на уплотнитель — т.н. «зимний» режим, наоборот, от уплотнителя — т.н. «летний» режим. Рекомендуется для снижения износа фурнитуры в теплое вемя года использовать летний режим, а при использовании зимой зимнего режима, переводить обратно при наступлении теплого времени года.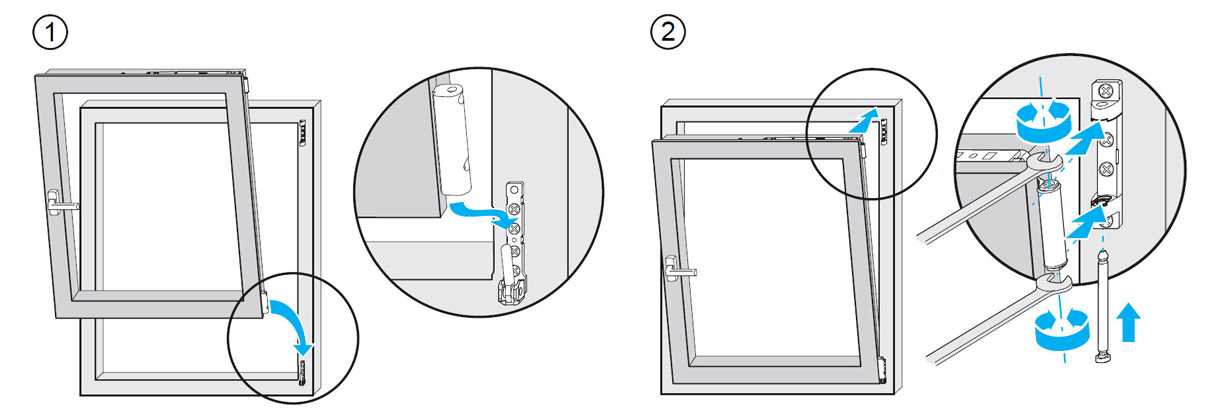 При постановке зимнего режима нагрузка на фурнитуру повышается, оконная ручка поворачивается туже. Цапфы могут иметь различную форму и различный паз под регулировку. Чаще всего это паз под шестигранник с тем же размером, что и винты петель, но встречается и несоответствие. На некоторых системах цапфы не имеют паза, и их вращение производится при помощи пассатиж либо подобного инструмента (фурнитура ‘МАСО’).
При постановке зимнего режима нагрузка на фурнитуру повышается, оконная ручка поворачивается туже. Цапфы могут иметь различную форму и различный паз под регулировку. Чаще всего это паз под шестигранник с тем же размером, что и винты петель, но встречается и несоответствие. На некоторых системах цапфы не имеют паза, и их вращение производится при помощи пассатиж либо подобного инструмента (фурнитура ‘МАСО’).
САМЫЕ РАСПРОСТРАНЕННЫЕ СЛУЧАИ
Устранение нарушения работы створок
ВНИМАНИЕ!
Важно помнить, что все действия с оконной ручкой должны производиться при полностью прижатой к раме окна створке. В противном случае возможно возникновение такого казуса, как т.н. «двойное открывание».
«Двойное открывание»
Створка окна находится отновременно и открытой и откинутой. Такое ощущение, что створка «вывалилась из верхней петли» и «вот-вот рухнет на пол». Происходит это вследствие неправильного использования окна. Т.е., створка была открыта, Вы решили ее откинуть (или наоборот), но не до конца прижали ее к раме. Либо это произошло по причине сильного проседания створки.
Либо это произошло по причине сильного проседания створки.
В фурнитуре створки часто имеется специальное устройство, которое называется блокиратор ошибочного действия или микролифт. Это устройство предназначено для предотвращения поворота оконной ручки при открытой/откинутой створке, а также служит для приподнимания створки для защиты от провисания. Когда створка закрыта, язычок микролифта прижат, и блокиратор выключен. Если повернуть ручку при не до конца прижатой створке, то язычок прижаться успевает, а фурнитура не успевает перейти следующее положение. Блокиратор-микролифт — это очень полезная деталь в оконной фурнитуре, но совсем необязательная. Она лишь призвана повысить комфорт и повысить срок службы окна. Именно поэтому довольно часто такая опция не входит в базовую комплектацию окна, и, соответственно, Вы можете не встретить на своем окне такого устройства. Чтобы устранить этот дефект, не обязательно вызывать мастера. Вы сможете все исправить сами за считанные секунды. Достаточно выполнить следующие действия:
1. Прижать одной рукой язычок микролифта параллельно уплотнителю. Другой рукой повернуть оконную ручку в положение «откинуто», не отпуская язычок микролифта. (Если ручка уже находится в этом положении, этот шаг пропускаем). Для створок, которые не оборудованы блокиратором ошибочного действия — просто перевести ручку в положение «Откинуто». Важно, чтобы в момент поворота ручки створка была близка к положению «Откинуто», т.е. верхний угол створки на петлевой стороне должен быть отведен от рамы.
Прижать одной рукой язычок микролифта параллельно уплотнителю. Другой рукой повернуть оконную ручку в положение «откинуто», не отпуская язычок микролифта. (Если ручка уже находится в этом положении, этот шаг пропускаем). Для створок, которые не оборудованы блокиратором ошибочного действия — просто перевести ручку в положение «Откинуто». Важно, чтобы в момент поворота ручки створка была близка к положению «Откинуто», т.е. верхний угол створки на петлевой стороне должен быть отведен от рамы.
2. Прижать рукой верхний угол створки с петлевой стороны.
Так же, как и в самом начале, прижать язычок микролифта и перевести ручку в положение «открыто». Для створок, которые не оборудованы блокиратором ошибочного действия — просто перевести ручку в положение «Открыто». Все должно работать. Если не помогло, значит створка просела вниз (бывает очень редко). Поднять створку, вращая винт № 1.
Провисание створки
Это самый распространенный случай при рекламационных обращениях. Проявляется в затруднении закрывания створки, ввиду ее контакта с рамой. Т.е. «створка цепляет раму» или элементы фурнитуры, закрепленные на раме. Причиной может служить следующее (каждый пункт может повлечь содержание какого-либо другого пункта или нескольких. Все нижеописанные причины актуальны при сохранении статичности оконного проема.):
Проявляется в затруднении закрывания створки, ввиду ее контакта с рамой. Т.е. «створка цепляет раму» или элементы фурнитуры, закрепленные на раме. Причиной может служить следующее (каждый пункт может повлечь содержание какого-либо другого пункта или нескольких. Все нижеописанные причины актуальны при сохранении статичности оконного проема.):
- Нарушение правил сборки окна. Т.е. габаритные размеры створки не сочетаются с габаритными размерами рамы. Проверяется следующим образом: закройте створку и обведите ее карандашом по раме; откройте створку. Расстояние от края рамы до очерченной линии должно составлять 5-8 мм. Допускаются незначительные отклонения. Устранения можно добиться постоянными регулировками.
- «Эффект новой петли». Чаще всего проявляется в первые полгода эксплуатации. Нижняя петля створки одевается на штифт опоры петли. Петля имеет внутри пластиковый чулок для снижения шума при работе створки. Иногда новая петля имеет т.н. заусенец внутри пластикового чулка.
 Со временем этот заусенец притирается, и створка садится на место. Может проявляться в виде «двойного открывания» (см. случай № 1), контакта с рамой или с рамными деталями, оконная ручка не проворачивается до конца. Устраняется путем поднятия створки. Вращаем винт в нижней петле. При необходимости переставляем запорные пластины повыше/пониже.
Со временем этот заусенец притирается, и створка садится на место. Может проявляться в виде «двойного открывания» (см. случай № 1), контакта с рамой или с рамными деталями, оконная ручка не проворачивается до конца. Устраняется путем поднятия створки. Вращаем винт в нижней петле. При необходимости переставляем запорные пластины повыше/пониже. - Неправильная постановка стеклопакета в створке. Стеклопакет не расклинен. Это приводит к тому, что створка принимает форму параллелограмма, нижний противоположный петле угол стремится вниз, и тем самым вызывает контакт с рамой. Устраняется вращением винта верхней петли в сторону петли (при незначительных отклонениях). При значительных — переустановка стеклопакета в створке. Значительность отклонений можно определить путем измерения диагоналей створки. При расхождении более чем на 10 мм скорее всего нужно будет вызывать специалиста для расклинивания стеклопакета.
- Некачественный профиль, фурнитура. Использование некачественных материалов при сборке может привести к повышенному износу. Иными словами профиль и фурнитура могут «вытягиваться» со временем. Постоянные регулировки, до крайнего положения винта № 3, после чего не устраняется.
 Со временем этот заусенец притирается, и створка садится на место. Может проявляться в виде «двойного открывания» (см. случай № 1), контакта с рамой или с рамными деталями, оконная ручка не проворачивается до конца. Устраняется путем поднятия створки. Вращаем винт в нижней петле. При необходимости переставляем запорные пластины повыше/пониже.
Со временем этот заусенец притирается, и створка садится на место. Может проявляться в виде «двойного открывания» (см. случай № 1), контакта с рамой или с рамными деталями, оконная ручка не проворачивается до конца. Устраняется путем поднятия створки. Вращаем винт в нижней петле. При необходимости переставляем запорные пластины повыше/пониже. Иными словами профиль и фурнитура могут «вытягиваться» со временем. Постоянные регулировки, до крайнего положения винта № 3, после чего не устраняется.
Иными словами профиль и фурнитура могут «вытягиваться» со временем. Постоянные регулировки, до крайнего положения винта № 3, после чего не устраняется.Поддувание из-под створки
Створка современного окна позволяет избежать сильного продувания, и их уже не нужно заклеивать на зиму, как старые рамы. Тем не менее, створки пластикового окна также пропускают воздух (имеют свой коэффициент воздухопроницаемости), но в значительно меньших количествах. Рассмотрим случаи, когда воздухопроницаемость створки вызывает дискомфорт.
- Из-под уплотнителя сильно дует в каком-либо одном месте или по всему периметру. Дует иногда «с завываниями» при не сильном ветре на улице. Причина в отсутствии должного прижима ввиду отсутствия или несоответствия запорных пластин, их неправильной установки, неправильной предыдущей регулировки створки либо габаритные размеры створки не сочетаются с габаритными размерами рамы. Устраняется путем правильной установки запорных пластин нужной серии, вращением винта опоры нижней петли в сторону проблемных запорных пластин. В случае с «негабаритами» — не устраняется.
- То же, что в п.1, но никаких нарушений сборки нет. Причиной может быть т.н. «гипервентиляция», — являение, при котором подсос воздуха с улицы осуществляется разностью давления воздуха. Может проявляться достаточно сильно, вплоть до деформации рам. Устраняется при помощи обеспечения нормального притока воздуха посредством стеновых клапанов, рекуперационных установок и проч.
- Из-под уплотнителя «тянет» холодный воздух. Особенно в районе нижней петли и нижней горизонтальной стороны створки. Сильного продувания нет, но ощущается холодный ток воздуха. В зависимости от температуры теплоносителя и правильности постановки подоконной доски ощущение «продувания» может либо усиливаться либо ослабляться. Также это ощущение физиологически может быть различным для восприятия у разных людей. Именно для этого и существует возможность перевода цапф в т.н. «зимний» режим.
- То же, что в п.1 или 2, т.н. «зимний» режим не помогает. Возможно нарушена эластичность створочных уплотнителей. Резиновые уплотнители изначально имеют специальную силиконовую смазку, которая со временем «съедается» пылью, стирается. Поэтому необходимо ухаживать за окном, периодически смазывать и протирать уплотнители. При необходимости заменить уплотнитель.
 В случае с «негабаритами» — не устраняется.
В случае с «негабаритами» — не устраняется. Резиновые уплотнители изначально имеют специальную силиконовую смазку, которая со временем «съедается» пылью, стирается. Поэтому необходимо ухаживать за окном, периодически смазывать и протирать уплотнители. При необходимости заменить уплотнитель.
Резиновые уплотнители изначально имеют специальную силиконовую смазку, которая со временем «съедается» пылью, стирается. Поэтому необходимо ухаживать за окном, периодически смазывать и протирать уплотнители. При необходимости заменить уплотнитель.Оконная ручка шатается, ощущение ее «хлипкости»
Оконная ручка закрепляется к створке при помощи двух винтов. Иногда, в процессе эксплуатации затяжка этих винтов ослабевает. Устраняется все очень просто.
- Сдвинуть крышку оконной ручки в сторону.
- Затянуть крестовой отверткой винты. Важно помнить, что чрезмерное усилие при затягивании винтов может привести к поломке оконной ручки.
Регулировка площади листьев как оптимальная стратегия адаптации к засухе
- Манцони, С.
- Бейер, Ф.
- Томпсон, С.Э.
- Вико, Г.
- Вейх, М.
;
;
;

;
Аннотация
Фенология листьев играет важную роль в обмене массой и энергией между землей и атмосферой. Много работы было сосредоточено на фенологических реакциях на свет и температуру, но меньше на изменениях площади листьев в засушливые периоды. Поскольку продолжительность засух, как ожидается, увеличится в будущем климате как в сезонно-сухой, так и в мезической среде, крайне важно (i) предсказать фенологические изменения, связанные с засухой, и (ii) разработать физиологически обоснованные модели динамики площади листьев в течение засушливые периоды. Было предложено несколько критериев оптимизации для моделирования регулирования площади листьев по мере уменьшения влажности почвы. Некоторые теории основаны на балансе углерода (C) в растениях, предполагая, что площадь листьев уменьшится, когда мгновенные чистые скорости фотосинтеза станут отрицательными (что эквивалентно максимизации кумулятивного прироста C). Другие теории основаны на гидравлических принципах, предполагая, что площадь листа должна регулироваться либо для поддержания постоянного водного потенциала листа (изогидрическое поведение), либо для предотвращения отрицательного воздействия водного потенциала листа на фотосинтез (т. Е. Минимизации водного стресса). Фенология вечнозеленых листьев рассматривается как контрольный случай. Объединив эти теории в единую структуру, мы количественно определяем влияние фенологической стратегии и воздействия климата на чистый прирост углерода в течение всего вегетационного периода. Путем учета затрат углерода на смывание листьев и выгод, связанных с фотосинтезом листьев, этот показатель оценивает эффективность различных фенологических стратегий при различных климатических сценариях. Вечнозеленые виды предпочтительны только тогда, когда засушливый период относительно короткий, поскольку они могут использовать большую часть вегетационного периода и несут затраты на уход за листьями только в течение короткого засушливого периода.
Другие теории основаны на гидравлических принципах, предполагая, что площадь листа должна регулироваться либо для поддержания постоянного водного потенциала листа (изогидрическое поведение), либо для предотвращения отрицательного воздействия водного потенциала листа на фотосинтез (т. Е. Минимизации водного стресса). Фенология вечнозеленых листьев рассматривается как контрольный случай. Объединив эти теории в единую структуру, мы количественно определяем влияние фенологической стратегии и воздействия климата на чистый прирост углерода в течение всего вегетационного периода. Путем учета затрат углерода на смывание листьев и выгод, связанных с фотосинтезом листьев, этот показатель оценивает эффективность различных фенологических стратегий при различных климатических сценариях. Вечнозеленые виды предпочтительны только тогда, когда засушливый период относительно короткий, поскольку они могут использовать большую часть вегетационного периода и несут затраты на уход за листьями только в течение короткого засушливого периода. Напротив, лиственные виды, которые снижают затраты на содержание за счет потери листьев, имеют преимущество в более сухом климате. Более того, среди засушливых лиственных видов изогидрическое поведение приводит к наименьшему приросту углерода. Постепенная потеря листьев, чтобы поддерживать чистое поглощение углерода равным нулю в самый засушливый период вегетационного периода, обеспечивает наибольший прирост. Поскольку все эти стратегии определяются на основе часто моделируемых величин, их можно реализовать в моделях экосистем в зависимости от функционального типа растения и климата.
Напротив, лиственные виды, которые снижают затраты на содержание за счет потери листьев, имеют преимущество в более сухом климате. Более того, среди засушливых лиственных видов изогидрическое поведение приводит к наименьшему приросту углерода. Постепенная потеря листьев, чтобы поддерживать чистое поглощение углерода равным нулю в самый засушливый период вегетационного периода, обеспечивает наибольший прирост. Поскольку все эти стратегии определяются на основе часто моделируемых величин, их можно реализовать в моделях экосистем в зависимости от функционального типа растения и климата.
- Публикация:
Тезисы осенней встречи AGU
- Дата публикации:
- Декабрь 2014 г.
- Биб-код:
2014АГУФМ.
Б23Ф0286М- Ключевые слова:
- 0414 Биогеохимические циклы;
- процесса;
- и моделирование;
- БИОЛОГИЧЕСКИЕ НАУКИ;
- 0470 Питательные вещества и круговорот питательных веществ;
- БИОЛОГИЧЕСКИЕ НАУКИ;
- 0476 Экология растений;
- БИОЛОГИЧЕСКИЕ НАУКИ;
- 1615 Биогеохимические циклы;
- процесса;
- и моделирование;
- ГЛОБАЛЬНОЕ ИЗМЕНЕНИЕ
 Б23Ф0286М
Б23Ф0286МКорректировка анатомических и гидравлических характеристик листьев по вертикальным профилям полога молодых лиственных насаждений
Аасамаа К. , Сыбер А. (2012) Светочувствительность гидравлической проводимости побегов у пяти лиственных пород умеренного пояса. Функц Биол 39:661–669
, Сыбер А. (2012) Светочувствительность гидравлической проводимости побегов у пяти лиственных пород умеренного пояса. Функц Биол 39:661–669
CAS
пабмед
Google ученый
Aasamaa K, Niinemets Ü, Sõber A (2005) Гидравлическая проводимость листа в зависимости от анатомических и функциональных признаков в онтогенезе листа Populus tremula . Tree Physiol 25:1409–1418
PubMed
Google ученый
Аасамаа К., Кыйвик К., Куппер П., Сыбер А. (2014) Среда роста определяет светочувствительность гидравлической проводимости побегов. Эколь Рез 29: 143–151
Google ученый
Abrams MD, Kubiske ME (1990) Структурные характеристики листа 31 породы лиственных и хвойных деревьев в центральном Висконсине: влияние светового режима и степени теневыносливости. Forest Ecol Manag 31:245–253
Google ученый
Al Afas N, Pellis A, Niinemets Ü, Ceulemans R (2005) Выращивание и производство порослевой культуры тополя с коротким оборотом. II. Клоновые и межгодовые различия в характеристиках листа и черешка и индексе листовой поверхности. Биомасса Биоэнергетика 28:536–547
II. Клоновые и межгодовые различия в характеристиках листа и черешка и индексе листовой поверхности. Биомасса Биоэнергетика 28:536–547
Google ученый
Блондер Б., Виолле С., Энквист Б.Дж. (2013) Оценка причин и масштабов спектра экономики листьев с использованием сетей жилкования Populus tremuloides . J Ecol 101:981–989
Google ученый
Boonman CCF, Бенитес-Лопес А., Шиппер А.М. и др. (2020) Оценка надежности прогнозируемых распределений признаков растений в глобальном масштабе. Глоб Эколь Биогеогр 29:1034–1051
PubMed
ПабМед Центральный
Google ученый
Бреда Н., Судани К., Бергонзини Дж.-К. (2002) Mesure de l’indice foliaire en forêt. Ecofor, Париж
Google ученый
Бродрибб Т.Дж., Филд Т.С., Джордан Г.Дж. (2007) Максимальная скорость фотосинтеза листьев и жилкование связаны гидравликой.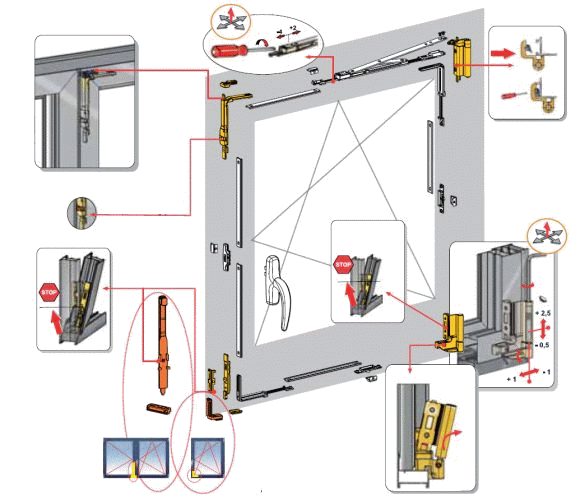 Plant Physiol 144:1890–1898
Plant Physiol 144:1890–1898
CAS
пабмед
ПабМед Центральный
Google ученый
Бакли Т.Н., Грейс П.Дж., Скоффони С., Сак Л. (2015) Как анатомия листа влияет на перенос воды за пределы ксилемы? Plant Physiol 168:1616–1635
CAS
пабмед
ПабМед Центральный
Google ученый
Кэмпбелл Г.С., Норман Дж.М. (1989) Описание и измерение структуры растительного покрова. В: Рассел Г., Маршалл Б., Джарвис П.Г. (ред.) Навесы растений: их рост, форма и функции общества для экспериментальной биологии. Серия семинаров 31. Издательство Кембриджского университета, Кембридж, стр. 1–19.
Google ученый
Каринс Мерфи М.Р., Джордан Г.Дж., Бродрибб Т.Дж. (2014) Акклиматизация к влажности изменяет связь между размером листа и плотностью жилок и устьиц. Plant Cell Environment 37:124–131
CAS
пабмед
Google ученый
Каринс Мерфи М. Р., Джордан Г.Дж., Бродрибб Т.Дж. (2017) Папоротники в меньшей степени зависят от пассивного разбавления за счет расширения клеток для координации жилок листа и расстояния между устьицами, чем покрытосеменные. ПЛОС ОДИН 12(9):e0185648
Р., Джордан Г.Дж., Бродрибб Т.Дж. (2017) Папоротники в меньшей степени зависят от пассивного разбавления за счет расширения клеток для координации жилок листа и расстояния между устьицами, чем покрытосеменные. ПЛОС ОДИН 12(9):e0185648
ПабМед
ПабМед Центральный
Google ученый
Casella E, Ceulemans R (2002) Пространственное распределение морфологических и физиологических характеристик листьев в зависимости от локального радиационного режима в кронах 3-летних клонов Populus в порослевой культуре. Tree Physiol 22:1277–1288
CAS
пабмед
Google ученый
Caudullo G, de Rigo D (2016) Populus tremula в Европе: распространение, среда обитания, использование и угрозы. В: Сан-Мигель-Аянц Дж., Де Риго Д., Кодулло Г., Хьюстон Даррант Т., Маури А. (ред.) Европейский атлас видов лесных деревьев. Издательское бюро ЕС, Люксембург, стр. 138–139
Google ученый
Чаздон Р.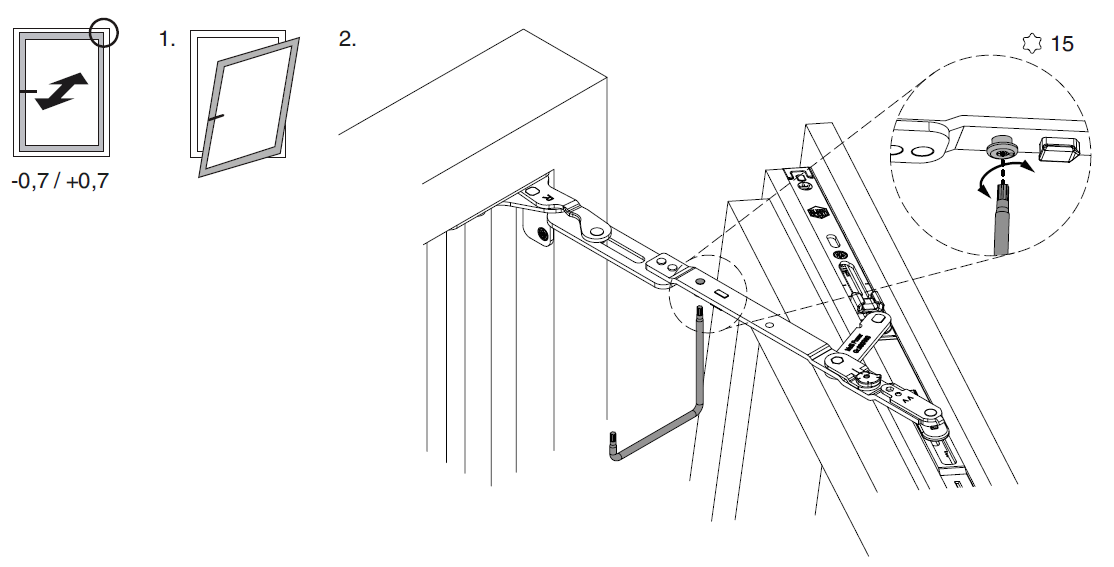 Л., Кауфманн С. (1993) Пластичность анатомии листьев двух кустарников тропических лесов в связи с фотосинтетической акклиматизацией к свету. Функция Экол 7: 385–394
Л., Кауфманн С. (1993) Пластичность анатомии листьев двух кустарников тропических лесов в связи с фотосинтетической акклиматизацией к свету. Функция Экол 7: 385–394
Google ученый
Кобл А.П., Кавалери М.А. (2014) Свет управляет вертикальными градиентами морфологии листьев в лесу сахарного клена ( Acer saccharum ). Tree Physiol 34: 146–158
CAS
пабмед
Google ученый
Кобл А.П., Кавалери М.А. (2015) Световая акклиматизация оптимизирует функциональные характеристики листа, несмотря на ограничения, связанные с высотой, в эксперименте по затенению кроны. Экология 177:1131–1143
ПабМед
Google ученый
Кусо Л.Л., Фернандес Р.Дж. (2012) Фенотипическая пластичность как показатель засухоустойчивости у трех патагонских степных трав. Энн Бот 110:849–857
CAS
пабмед
ПабМед Центральный
Google ученый
da Silveira TI, Boeger MRT, Maranho LT, de Melo JC, Soffiatti P (2015) Функциональные признаки листьев 57 древесных видов леса Араукария, Южная Бразилия.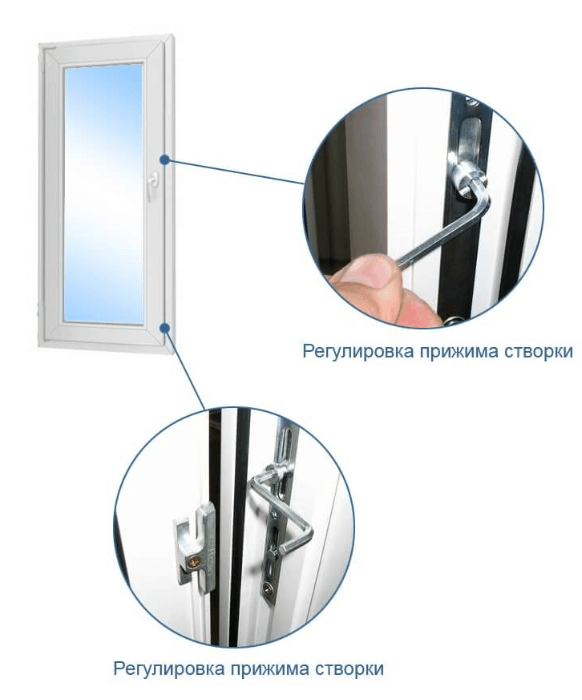 Браз Джей Бот 38: 357–366
Браз Джей Бот 38: 357–366
Google ученый
Дитрих Л., Хох Г., Кахмен А., Кёрнер К. (2018) Потеря половины проводящей площади практически не влияет на состояние воды взрослых деревьев. Научный представитель 8:15006
PubMed
ПабМед Центральный
Google ученый
Доу Г.Дж., Берри Дж.А., Бергманн Д.К. (2014) Физиологическая важность механизмов развития, обеспечивающих правильное расстояние между устьицами у Arabidopsis thaliana . Новый фитол 201:1205–1217
CAS
пабмед
Google ученый
Du Q, Liu T, Jiao X, Song X, Zhang J, Li J (2019) Анатомические адаптации листьев играют центральную роль в фотосинтетической акклиматизации к влажности. J Exp Bot 70: 4949–4961
CAS
пабмед
Google ученый
Дюран М., Брендель О., Буре С. , Ле Тьек Д. (2019)Измененная динамика устьиц, вызванная изменениями освещенности и дефицита давления пара в условиях засухи: влияние на эффективность транспирации всего растения генотипов тополя. Новый фитол 222:1789–1802
, Ле Тьек Д. (2019)Измененная динамика устьиц, вызванная изменениями освещенности и дефицита давления пара в условиях засухи: влияние на эффективность транспирации всего растения генотипов тополя. Новый фитол 222:1789–1802
КАС
пабмед
Google ученый
Eensalu E, Kupper P, Sellin A, Rahi M, Sõber A, Kull O (2008) Работают ли устьица с одним и тем же относительным диапазоном раскрытия вдоль профиля кроны Betula pendula ? Funct Plant Biol 35:103–110
PubMed
Google ученый
Fichot R, Chamaillard S, Depardieu C et al (2011) Гидравлическая эффективность и координация с устойчивостью ксилемы к кавитации, функцией листьев и показателями роста среди восьми не связанных между собой Populus deltoides × Populus nigra гибриды. J Exp Bot 62: 2093–2106
CAS
пабмед
Google ученый
Флорес-Морено Х. , Фазаели Ф., Банерджи А. и др. (2019) Надежность связей признаков в зависимости от градиентов окружающей среды и форм роста. Глоб Эколь Биогеогр 28:1806–1826
, Фазаели Ф., Банерджи А. и др. (2019) Надежность связей признаков в зависимости от градиентов окружающей среды и форм роста. Глоб Эколь Биогеогр 28:1806–1826
Google ученый
Franks PJ, Farquhar GD (2001) Влияние экзогенной абсцизовой кислоты на развитие устьиц, устьичную механику и газообмен листьев у Tradescantia virginiana . Plant Physiol 125:935–942
CAS
пабмед
ПабМед Центральный
Google ученый
Franks PJ, Drake PL, Beerling DJ (2009) Пластичность максимальной устьичной проводимости, ограниченная отрицательной корреляцией между размером устьиц и их плотностью: анализ с использованием Eucalyptus globulus . Plant Cell Environ 32:1737–1748
PubMed
Google ученый
Gamage HK, Ashton MS, Singhakumara BMP (2003) Структура листьев Syzygium spp. (Myrtaceae) в связи с близостью к участкам в тропических лесах. Бот Дж. Линн Сок 141: 365–377
Бот Дж. Линн Сок 141: 365–377
Google ученый
Hale SE, Edwards C (2002) Сравнение пленочных и цифровых полусферических фотографий в широком диапазоне плотностей кроны деревьев. Agric For Meteorol 112:51–56
Hanba YT, Kogami H, Terashima I (2002) Влияние освещения роста на анатомию листа и фотосинтез в Acer видов, отличающихся светолюбивостью. Окружающая среда клеток растений 25:1021–1030
Google ученый
Hansen EA (1991) Урожайность древесной биомассы тополя: взгляд в будущее. Биомасса Биоэнергия 1:1–7
Google ученый
Хараяма Х., Китао М., Агатоклеус Э., Исида А. (2019) Влияние закупорки основных вен и ингибирования аквапорином на гидравлику листа и устьичную проводимость. Proc R Soc B 286: 20190799
КАС
пабмед
ПабМед Центральный
Google ученый
Jaagus J, Mändla K (2014) Сценарии изменения климата для Эстонии на основе климатических моделей из Четвертого оценочного доклада МГЭИК.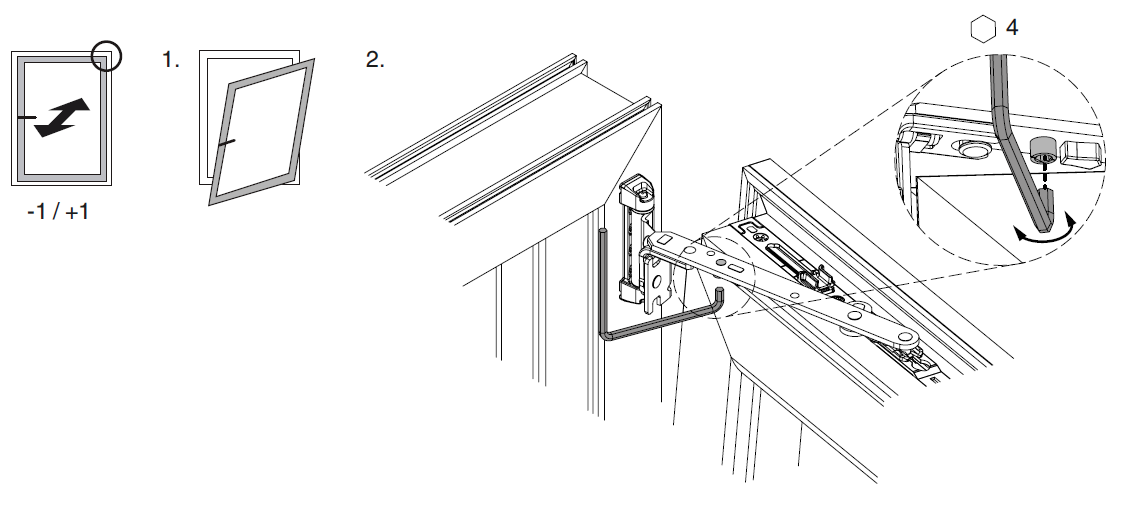 Эстонский J Earth Sci 63:166–180
Эстонский J Earth Sci 63:166–180
Google ученый
Джейкоб Д., Петерсен Дж., Эггерт Б. и др. (2014) EURO-CORDEX: новые прогнозы изменения климата с высоким разрешением для исследований воздействия на Европу. Reg Environ Change 14: 563–578
Google ученый
Ясинская А.К., Альбер М., Туллус А., Рахи М., Селлин А. (2015) Влияние повышенной атмосферной влажности на анатомические и гидравлические характеристики ксилемы гибридной осины. Funct Plant Biol 42:565–578
Google ученый
Йоханссон Т. (2013) Производство биомассы гибридной осины, растущей на бывших сельскохозяйственных угодьях в Швеции. J For Res 24:237–246
Кангур О., Степ К., Шрил Дж., фон дер Кроне Дж., Селлин А. (2021) Изменение ночной проводимости устьиц и развитие предрассветного неравновесия между потенциалами воды в почве и листьях у девяти видов лиственных деревьев умеренного пояса. Funct Plant Biol 48:483–492
Funct Plant Biol 48:483–492
CAS
пабмед
Google ученый
Колб К.Дж., Сперри Дж.С. (1999) Различия в адаптации к засухе между подвидами полыни ( Artemisia tridentata ). Экология 80:2373–2384
Google ученый
Кумагаи Т., Кураджи К., Ногучи Х., Танака Ю., Танака К., Судзуки М. (2001) Вертикальные профили факторов окружающей среды в тропических лесах, Национальный парк Ламбир-Хиллз, Саравак, Малайзия. J For Res 6:257–264
Kupper P, Sõber J, Sellin A et al (2011) Экспериментальная установка для управления влажностью свободного воздуха (FAHM) может изменять поток воды через крон лиственных деревьев. Бот Environ Exp 72: 432–438
Google ученый
Куппер П., Иванова Х., Сыбер А., Рохула-Окунев Г., Селлин А. (2018) Ночные и дневные водные отношения у пяти быстрорастущих пород деревьев: влияние экологических и эндогенных переменных. Экогидрология 11:e1927
Экогидрология 11:e1927
Google ученый
Lambers H, Poorter H (1992) Врожденные различия в скорости роста между высшими растениями: поиск физиологических причин и экологических последствий. Adv Ecol Res 23: 87–261
Google ученый
Lambers H, Chapin FS III, Pons TL (2006) Физиологическая экология растений. Спрингер, Нью-Йорк
Google ученый
Легнер Н., Флек С., Лойшнер С. (2013) Акклиматизация при слабом освещении пяти видов широколиственных деревьев умеренного пояса с разным сукцессионным статусом: значение тенистого навеса. Ann For Sci 70:557–570
Льюис А.М., Буз Э.Р. (1995) Оценка объемной скорости потока через ксилемные каналы. Am J Bot 82: 1112–1116
Google ученый
Li L, McCormack ML, Ma C et al (2015) Экономика листьев и гидравлические характеристики разделены в пяти богатых видами тропических и субтропических лесах. Ecol Lett 18:899–906
Ecol Lett 18:899–906
CAS
пабмед
Google ученый
Ли Ф, Маккалох К.А., Сун С., Бао В. (2018) Связь гидравлических свойств листьев, скорости фотосинтеза и продолжительности жизни листьев у ксерофитных видов: проверка глобальных гипотез. Ам Дж. Бот 105: 1858–1868
КАС
пабмед
Google ученый
Lihavainen J, Ahonen V, Keski-Saari S, Sõber A, Oksanen E, Keinänen M (2017) Низкий дефицит давления пара снижает плотность железистых трихом и изменяет химический состав кутикулярного воска в листьях березы повислой. Tree Physiol 37:1166–1181
CAS
пабмед
Google ученый
Мандре М., Туллус Х., Тамм Ю (1998) Распределение углеводов и биомассы листьев Populus tremula L. полога. Деревья 12:160–166
Google ученый
Мартинес-Вилальта Дж. , Гарсия-Форнер Н. (2017) Регулирование водного потенциала, поведение устьиц и гидравлический перенос в условиях засухи: деконструкция изо/анизогидрической концепции. Plant Cell Environ 40:962–976
, Гарсия-Форнер Н. (2017) Регулирование водного потенциала, поведение устьиц и гидравлический перенос в условиях засухи: деконструкция изо/анизогидрической концепции. Plant Cell Environ 40:962–976
PubMed
Google ученый
МакКаун А.Д., Кочард Х., Сак Л. (2010) Расшифровка гидравлики листа с помощью пространственно явной модели: принципы архитектуры жилкования и последствия для ее эволюции. Am Nat 175:447–460
PubMed
Google ученый
Mediavilla S, Martín I, Escudero A (2020) Признаки жилок и устьиц на листьях трех совместно встречающихся видов Quercus , различающихся продолжительностью жизни листьев. Eur J For Res 139: 829–840
Мессье Дж., Макгилл Б.Дж., Энквист Б.Дж., Лехович М.Дж. (2017) Изменение характеристик и интеграция в разных масштабах: присутствует ли экономический спектр листьев в локальных масштабах? Экография 40:685–697
Google ученый
Monclus R, Dreyer E, Villar M et al (2006) Влияние засухи на продуктивность и эффективность использования воды у 29 генотипов Populus deltoides × Populus nigra . Новый фитол 169: 765–777
Новый фитол 169: 765–777
ПабМед
Google ученый
Monsi M, Saeki T (2005) О факторе света в растительных сообществах и его важности для производства вещества. Энн Бот 95: 549–567
PubMed
ПабМед Центральный
Google ученый
Нардини А., Педа Г., Саллео С. (2012) Альтернативные методы масштабирования гидравлической проводимости листьев предлагают новое понимание структурно-функциональных отношений солнечных и теневых листьев. Функц Биол 39: 394–401
PubMed
Google ученый
Нардини А., Ыунапуу-Пикас Э., Сави Т. (2014) Чем меньше, тем лучше: гидравлическая проводимость листьев и уязвимость к засухе коррелируют с размером листьев и плотностью жилкования для четырех генотипов Coffea arabica . Funct Plant Biol 41:972–982
PubMed
Google ученый
Niinemets Ü (2001) Глобальный климатический контроль сухой массы листьев на единицу площади, плотности и толщины деревьев и кустарников. Экология 82: 453–469
Экология 82: 453–469
Google ученый
Niinemets Ü, Kull O, Tenhunen JD (1999) Изменчивость морфологии листьев и химического состава в зависимости от светового окружения кроны сосуществующих лиственных деревьев. Int J Plant Sci 160: 837–848
CAS
пабмед
Google ученый
Ниинеметс Ю, Эллсворт Д.С., Лукьянова А., Тобиас М. (2001) Плодородие участка и морфологическая и фотосинтетическая акклиматизация Pinus sylvestris Хвоя на свет. Tree Physiol 21:1231–1244
CAS
пабмед
Google ученый
Ниинеметс Ю, Кинан Т.Ф., Халлик Л. (2015) Всемирный анализ изменений структурных, химических и физиологических признаков листьев внутри полога по функциональным типам растений. Новый фитол 205:973–993
CAS
пабмед
Google ученый
Огучи Р.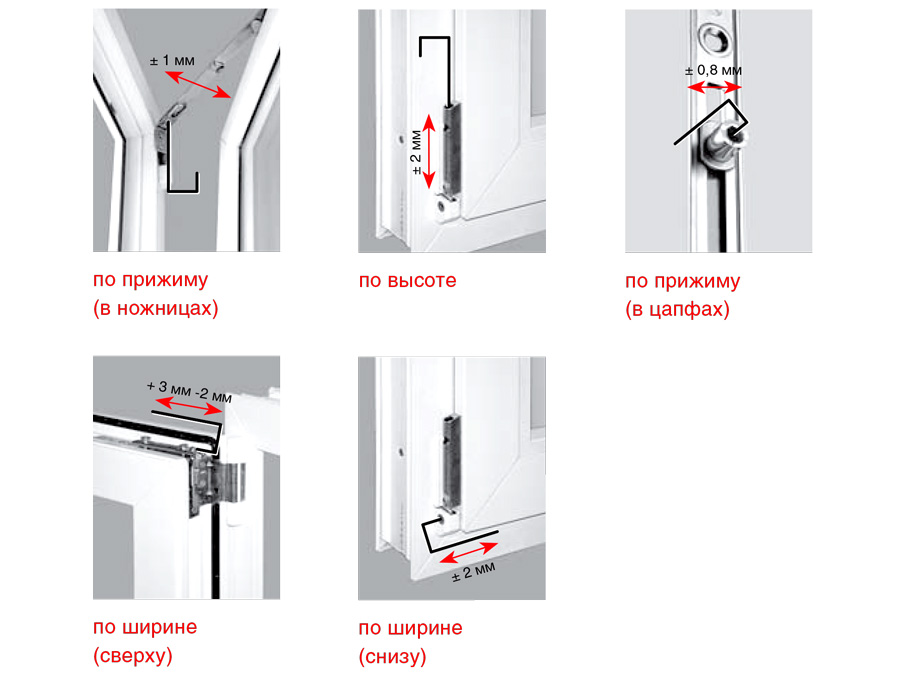 , Хикосака К., Хиросе Т. (2005) Анатомия листа как ограничение фотосинтетической акклиматизации: дифференциальная реакция анатомии листа на увеличение освещенности при росте у трех лиственных деревьев. Окружающая среда растительной клетки 28: 916–927
, Хикосака К., Хиросе Т. (2005) Анатомия листа как ограничение фотосинтетической акклиматизации: дифференциальная реакция анатомии листа на увеличение освещенности при росте у трех лиственных деревьев. Окружающая среда растительной клетки 28: 916–927
Google ученый
Õunapuu E, Sellin A (2013) Ежедневная динамика гидравлической проводимости листьев и почвы к ветвям березы повислой ( Betula pendula ), измеренная на месте. Plant Physiol Biochem 68:104–110
PubMed
Google ученый
Poorter H, Niinemets Ü, Poorter L, Wright IJ, Villar R (2009) Причины и последствия изменения массы листа на единицу площади (LMA): метаанализ. Новый Фитол 182: 565–588
ПабМед
Google ученый
Пуртер Х., Никлас К.Дж., Райх П.Б., Олексин Дж., Пут П., Моммер Л. (2012) Распределение биомассы по листьям, стеблям и корням: мета-анализ межвидовой изменчивости и контроля окружающей среды.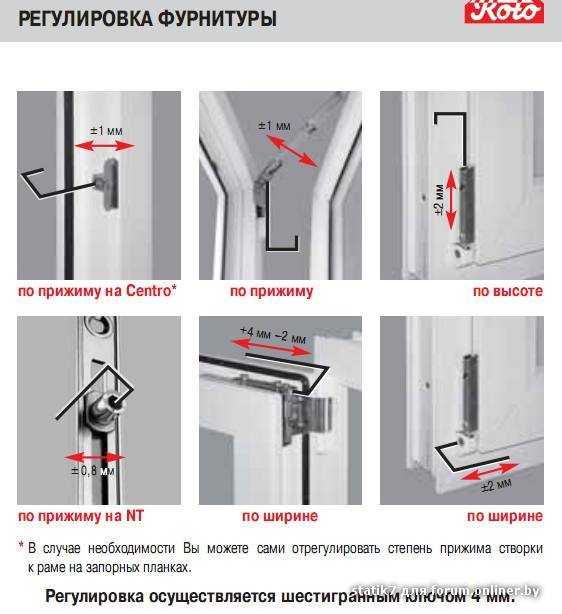 Новый фитол 193:30–50
Новый фитол 193:30–50
CAS
пабмед
Google ученый
Пройетти П., Паллиотти А., Фамиани Ф. и др. (2000) Влияние положения листьев, плодов и наличия света на фотосинтез двух генотипов каштана. Наука Хортик 85: 63–73
КАС
Google ученый
Рузин С.Е. (1999) Микротехника и микроскопия растений. Издательство Оксфордского университета, Оксфорд
Google ученый
Sack L, Frole K (2006) Структурное разнообразие листьев связано с гидравлической способностью деревьев тропических лесов. Экология 87:483–491
PubMed
Google ученый
Sack L, Holbrook NM (2006) Гидравлика листа. Annu Rev Plant Biol 57:361–381
CAS
пабмед
Google ученый
Сак Л., Скоффони К. (2013) Жилкование листьев: структура, функция, развитие, эволюция, экология и применение в прошлом, настоящем и будущем.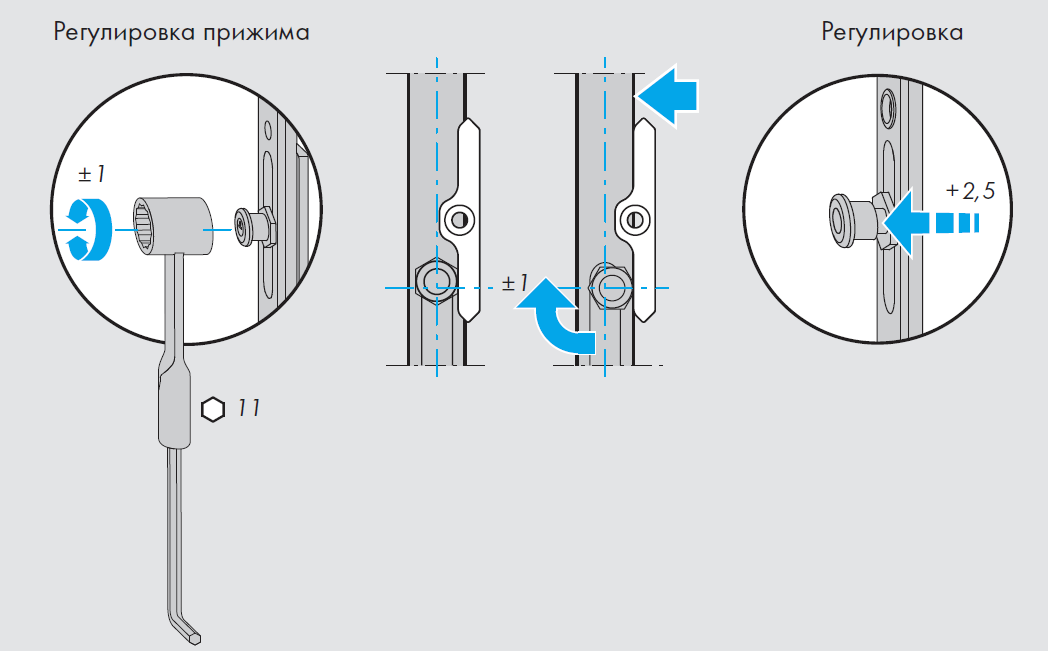 New Phytol 198:983–1000
New Phytol 198:983–1000
PubMed
Google ученый
Сак Л., Коуэн П.Д., Джайкумар Н., Холбрук Н.М. (2003) «Гидрология» листьев: координация структуры и функций древесных пород умеренного пояса. Окружающая среда растительных клеток 26:1343–1356
Google ученый
Сак Л., Мелчер П.Дж., Лю В.Х., Миддлтон Э., Парди Т. (2006) Насколько сильна пластичность листьев внутри полога лиственных деревьев умеренного пояса? Am J Bot 93: 829–839
PubMed
Google ученый
Сак Л., Скоффони С., Джон Г.П. и др. (2013) Как жилки листьев влияют на экономический спектр листьев во всем мире? Обзор и синтез. J Exp Bot 64: 4053–4080
КАС
пабмед
Google ученый
Сак Л., Скоффони С., Джонсон Д.М., Бакли Т.Н., Бродрибб Т.Дж. (2015) Анатомические детерминанты гидравлической функции листа. В: Hacke U (ed) Функциональная и экологическая анатомия ксилемы. Springer, Гейдельберг, стр. 255–271
В: Hacke U (ed) Функциональная и экологическая анатомия ксилемы. Springer, Гейдельберг, стр. 255–271
.
Google ученый
Скоффони С., Роулз М., Маккаун А., Кочард Х., Сак Л. (2011) Снижение гидравлической проводимости листа при обезвоживании: связь с размером листа и архитектурой жилкования. Физиол растений 156:832–843
КАС
пабмед
ПабМед Центральный
Google ученый
Скотт П. (2008) Физиология и поведение растений. Уайли, Чичестер
Google ученый
Селлин А., Куппер П. (2005) Влияние доступности света по сравнению с гидравлическими ограничениями на реакцию устьиц в кроне березы повислой. Экология 142:388–397
PubMed
Google ученый
Селлин А., Куппер П. (2006) Пространственное изменение отношения площади заболони к площади листьев и удельной площади листьев в кроне березы повислой. Деревья 20:311–319
Деревья 20:311–319
Google ученый
Селлин А., Куппер П. (2007) Влияние усиленного гидравлического питания листвы на реакцию устьиц мелколистной липы ( Tilia cordata Mill.). Eur J For Res 126:241–251
Sellin A, Õunapuu E, Kaurilind E, Alber M (2012) Изменчивость гидравлической проводимости листьев и побегов березы повислой в зависимости от размера. Деревья 26:821–831
Google ученый
Sellin A, Tullus A, Niglas A, Õunapuu E, Karusion A, Lõhmus K (2013) Изменение скорости роста, фотосинтетической способности, гидравлических свойств и других функциональных признаков у березы повислой под воздействием влажности ( Betula pendula ) . Ecol Res 28:523–535
CAS
Google ученый
Селлин А., Альбер М., Куппер П. (2017) Повышение влажности воздуха влияет на гидравлическую эффективность, но не на функциональную уязвимость ксилемы гибридной осины.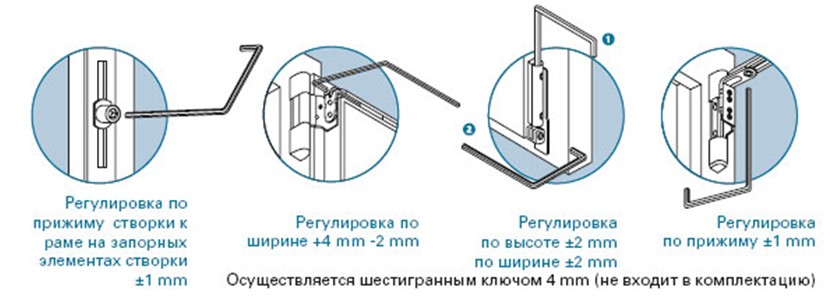 J Завод Физиол 219: 28–36
J Завод Физиол 219: 28–36
CAS
пабмед
Google ученый
Селлин А., Танеда Х., Альбер М. (2019) Структурная и гидравлическая регулировка листа березы повислой в зависимости от влажности воздуха и положения кроны ( Betula pendula ). J Plant Res 132: 369–381
CAS
пабмед
Google ученый
Смит Э.А., Коллетт С.Б., Бойнтон Т.А. и др. (2011) Вклад в развитие фенотипической изменчивости функциональных признаков листа в клонах дрожащей осины. Физиол дерева 31:68–77
ПабМед
Google ученый
Tullus A, Rytter L, Tullus T, Weih M, Tullus H (2012) Лесоводство с коротким оборотом гибридной осины ( Populus tremula L. × P. tremuloides Michx.) в Северной Европе. Scand J For Res 27:10–29
Tyree MT, Ewers FW (1991) Гидравлическая архитектура деревьев и других древесных растений.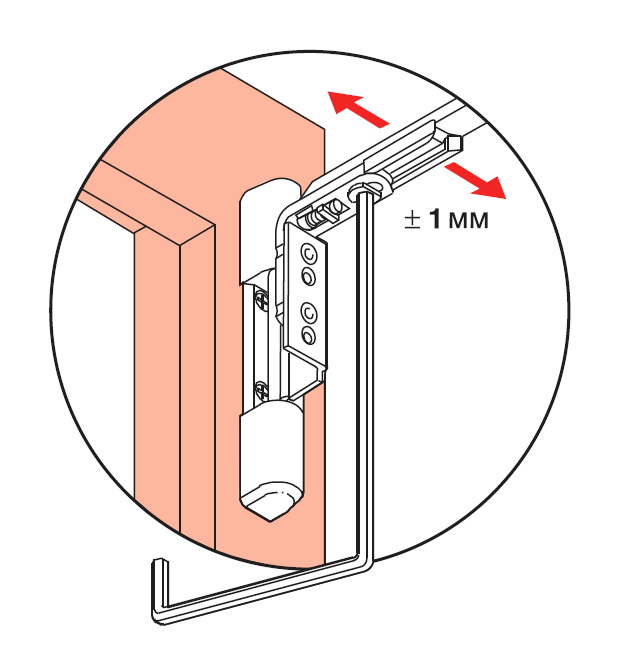 New Phytol 119:345–360
New Phytol 119:345–360
Witkowski ETF, Lamont BB (1991) Удельная масса листа путает плотность и толщину листа. Экология 88:486–493
CAS
пабмед
Google ученый
Райт И.Дж., Райх П.Б., Вестоби М. и др. (2004) Мировой спектр экономики листьев. Природа 428:821–827
CAS
пабмед
Google ученый
Xiong D, Flexas J, Yu T, Peng S, Huang J (2017) Анатомия листа опосредует координацию гидравлической проводимости листа и проводимости мезофилла по CO 2 в Орыза . Новый фитол 213: 572–583
CAS
пабмед
Google ученый
Yin Q, Wang L, Lei M et al (2018) Взаимосвязь между экономикой листьев и гидравлическими характеристиками древесных растений зависит от наличия воды. Sci Total Environ 621: 245–252
CAS
пабмед
Google ученый
Чжан Р, Ли И, Ни Ю, Гуй С, Лиан Дж, Е В (2019) Внутривидовая изменчивость функциональных признаков листа вдоль вертикального яруса в субтропическом вечнозеленом широколиственном лесу Дингушань.